광합성
지구상의 생물은 공기 중의 산소와 지상의 탄소화합물 때문에 살아갈 수 있는데, 이 2가지 모두 광합성에서 비롯된다. 광합성 과정은 지구가 탄생한 지 약 10억 년 뒤에 생겨난 것으로 추측되며 광합성의 시작은 생물세계가 출현할 수 있는 실마리를 제공했다. 인간의 입장에서 보았을 때 광합성은 단순한 식량뿐 아니라 석탄·석유와 같은 화석연료도 얻게 해주었다.

발견
광합성에 대한 연구는 1771년 영국의 화학자 조지프 프리스틀리가 시작했다. 프리스틀리는 밀폐된 용기 안에 식물이 있을 경우, 식물에서 어떤 물질(후에 산소로 밝혀짐)이 나와 공기가 제한되어 있는데도 초가 계속 탄다는 것을 발견했다.
1779년 네덜란드의 얀 잉겐호우스는 프리스틀리의 생각을 발전시켜, 가연성의 물질(산소)이 나오려면 식물이 빛을 받아야 하며 이때 식물의 녹색조직이 필요함을 밝혔다.
1782년 산소와 더불어 또다른 기체(고정되는 기체, 즉 이산화탄소)가 관여한다는 것이 알려졌고, 1804년 식물의 무게가 증가하는 것은 흡수한 탄소와 물 때문인 것으로 밝혀졌다. 1845년에는 광합성 과정중에 빛 에너지가 화학 에너지로 바뀐다는 것이 밝혀졌다.
전체 반응
화학 용어로 말하자면 광합성은 빛 에너지에 의한 산화환원(산화는 분자가 전자를 잃는 것이고 환원은 전자를 얻는 것을 말함)과정이라고 할 수 있다. 식물의 광합성에서는 빛 에너지를 사용해 물(H2O)이 산화하여 산소(O2), 수소 이온(H+), 전자 등이 발생한다.
이때 이산화탄소(CO2)가 대부분의 전자와 수소 이온을 받아 유기물질로 환원되며 그 주요산물은 녹말이나 설탕과 같은 탄수화물이다.
식물의 광합성을 통해 탄수화물(CH2O)이 만들어지는 반응은 다음과 같이 요약해서 나타낼 수 있다.
광합성의 전체반응은 빛에 의존하는 광화학 반응인 '명반응'과 유기 촉매인 효소에 의해 조절되는 화학반응인 '암반응'으로 나눌 수 있다.
명반응에서 빛에너지는 일련의 전자전달과정에 이용되어 궁극적으로 에너지가 풍부한 화합물인 아데노신삼인산(ATP)과 환원된 니코틴아미드아데닌디뉴클레오티드인산(NADPH) 을 만든다. 탄소고정과정인 암반응에서는 이 ATP와 NADPH를 이용하여 이산화탄소를 유기탄소화합물로 만든다.
황세균의 경우는 물 대신 황화수소(H2S)로부터 수소 이온을 만들고 산소 대신 황을 내놓는다.
녹색식물의 진화
현재의 녹색식물은 최초의 원시세포에서 진화해온 것으로 생각된다.
초기 지구에서는 전기 폭풍우, 태양에너지의 도움으로 물·암모니아·메탄과 같은 간단한 물질로부터 복잡한 분자들이 형성되고 최초의 원시세포가 이러한 분자들로부터 진화했을 것으로 생각하고 있다. 이런 과정에서 엽록소와 같은 색소를 지니게 된 원시세포가 빛 에너지를 사용할 수 있도록 진화했을 것이다(→ 채색).
20~30억 년 전 산소를 발생할 수 있었던 첫번째 세포는 남조류로 추측되는데, 이러한 작은 생물이 오랜 세월 동안 대기 중의 산소량을 증가시켜 호기성 생물을 나타나게 했을 것이라 생각된다.
남조류는 핵이나 엽록체와 같이 분막으로 싸인 세포 소기관이 없는 원핵생물이고 현재의 녹색식물은 이러한 소기관이 있는 진핵생물이기 때문에, 광합성을 했던 최초의 진핵생물은 비광합성 진핵생물이 남조류를 삼킨 후 진화한 홍조류일 것으로 추측하고 있다. 또한 오랜 세월이 지나는 동안 숙주세포 안의 남조류는 지금과 같은 엽록체로 진화했을 것이다.
엽록체의 구조와 구성요소
개요
식물에서 광합성의 모든 과정은 엽록체 안에서 일어난다. 이 세포 소기관의 기능에 대한 자세한 연구는 영국의 생물학자 로버트 힐이 시작했다. 1940년 힐은 깨진 세포에서 얻은 녹색 조각들에 빛과 전자수용체인 옥살산철을 주었을 때 산소가 발생하는 힐 반응을 발견했다. 1950년대에 미국의 생물학자 다니엘 아논은 전자수용체로 조효소 NADP를 사용하여 힐 반응뿐 아니라 에너지 저장물질인 ATP도 합성하는 식물세포 조각을 분리해냈다.
구조
엽록체는 2중막으로 되어 있으며 폭 2,500nm(1nm=10-9m), 길이 5,000nm의 타원형이다. 엽록체의 잘라진 부분을 전자현미경으로 조사하면 몇 가지 특징을 알 수 있다. 주요특징은 복잡한 막구조인 라멜라(lamellae)와 그것을 둘러싼 투명한 기질인 스트로마(stroma)를 들 수 있으며 그밖에 녹말 알갱이도 있다. 스트로마에는 암반응에 관련된 효소들이 녹아 있어서 탄소고정이 일어난다.
광합성에 필요한 모든 색소가 있는 라멜라는 10~15nm 두께의 납작한 모양으로 어떤 것은 엽록체 길이와 거의 맞먹는다. 라멜라를 가로로 잘라 보면, 가장자리가 서로 연결되어 틸라코이드(thylakoid)라는 속이 빈 납작한 원반 모양의 구조를 하고 있다. 고등식물에서 틸라코이드는 빽빽이 쌓여 그라나(grana)라는 구조를 만들며, 그라나와 그라나 사이는 긴 틸라코이드로써 연결되어 있다.
라멜라의 구성요소
① 색소:라멜라의 막에는 엽록소 a와 b가 단백질과 엽록소-단백질 복합체를 형성하고 있다. 이들은 빛에너지를 흡수해 엽록소 a분자에 전달해 주는데, 여기서 빛에너지가 화학에너지로 바뀌게 되며 엽록소 a가 빛에너지를 받아 흥분할 때 전자 하나를 내놓게 된다. 이런 특수한 엽록소 a분자들에는 2가지 유형이 있는데, 하나는 파장 684nm의 빛을 가장 잘 받아들이는 P680이고 다른 하나는 700nm의 빛을 가장 잘 받아들이는 P700이다.
엽록소말고도 카로틴이나 카로티노이드(홍당무의 색깔을 띠게 하는 색소) 등의 다른 색소도 빛을 흡수하나 이러한 색소가 흡수한 빛에너지는 다시 엽록소로 전달된 다음에야 화학에너지로 바뀐다.
② 단백질:엽록소-단백질 복합체뿐 아니라 그외에도 여러 가지 단백질들이 있다. 그중 중요한 것은 단백질조효소로서 전자를 전달하는 시토크롬(cytochrome)이다. 시토크롬에는 철원자가 든 색소가 있기 때문에 철원자의 산화·환원을 통해 전자를 전달할 수 있다. 그외에도 망간·철·구리가 포함된 라멜라 단백질체들이 있는데 이 단백질들의 금속원자는 광합성에서 중요한 촉매기능을 한다.
물이 분해되어 산소가 발생하는 데에는 망간이 작용하며, 명반응에서 물과 마지막 전자수용체 사이에서 전자를 전달하는 데에는 구리와 철을 지닌 단백질의 기능이 필요하다. 마지막 전자수용체인 페레독신(ferredoxin)은 철원자를 지닌 단백질이고 플라스토시아닌(plastocyanin)은 구리를 지닌 단백질인데 수용성이기 때문에 틸라코이드 공간 이동하면서 전자를 전달한다.
③ 퀴논(quinones):플라스토퀴논이라는 작은 분자들이 많이 있는데, 이들도 시토크롬과 마찬가지로 명반응의 여러 효소들 사이에서 전자를 전달하는 역할을 한다. 지질에 녹을 수 있기 때문에 막에서 확산을 통해 전자를 하나 또는 둘씩 운반한다.
명반응
빛의 흡수와 에너지 전이
빛을 받아들인 엽록소는 자신이 갖고 있는 전자 몇 개를 높은 에너지 수준으로 들뜨게 한다(→ 여기). 엽록소 a는 붉은색 빛(파장 약 680nm)을 받았을 때 들뜬 상태가 가장 낮다.
가장 낮게 들뜬 상태에 있는 엽록소 a분자의 전자가 원래의 에너지 상태인 바닥상태로 되돌아오면서 그만큼의 에너지를 내놓게 되는데, 이 에너지는 대부분 P680이나 P700과 같은 특별한 엽록소 a분자에서 다른 전자수용체로 전자를 전달하는 데 이용된다.
전자 전달 경로
전자가 물에서 이산화탄소로 이동하는 동안 2개의 명반응이 일어난다는, 1960년에 발표된 로버트 힐과 데렉 벤들의 이론이 널리 받아들여지고 있다.
세로축의 값은 산화·환원을 하면서 전자전달 경로에 관여하는 여러 가지 요소들의 상대적인 전위차를 볼트로 나타낸 것이다. 아래쪽으로 갈수록 전자를 빼앗으려는 성질(산화력)이 강하고 위쪽으로 갈수록 전자를 내주려는 성질(환원력)이 강하다.
명반응 Ⅱ에서 바닥상태의 P680이 들뜬상태(P680)가 되면 환원력이 매우 강해져서 엽록소와 비슷한 구조를 한 페오피틴(pheophytin)에 전자를 넘긴다. 역시 환원력이 강한 페오피틴은 다음 전자수용체인 QA와 QB라는 특수한 퀴논에 전자를 전달하고, 계속해서 전자는 플라스토퀴논풀(PQ pool), 철-황(Fe-S)단백질과 결부된 시토크롬 b와 f(Cytb와 Cytf)로 전달된다.
명반응 Ⅰ에서는 아직 알려지지 않은 첫번째 전자수용체 X가 들뜬 상태인 P700으로부터 전자를 받아서 철-황단백질에 넘겨주면, 전자는 작은 수용성 철-황단백질인 페레독신(Fd)을 거쳐 NADP+에 전달된다. 전자를 내놓고 바닥상태가 된 P680과 P700 은 양의 전하를 띠게 되며(P680+와 P700+), 강력한 산화력을 갖는다.
명반응 Ⅱ의 P680+는 망간-단백질복합체의 촉매작용으로 물 2분자로부터 4개의 전자를 얻어 P680으로 환원되는데 이때 알려지지 않은 전자운반체 Z를 거쳐서 전자를 받는다.
명반응 Ⅰ에서 P700+는 플라스토시아닌(PC)으로부터 전자를 받으며 PC는 PQ풀과 시토크롬b, 시토크롬f로부터 전자를 받는다. 전자가 물에서부터 여러 가지 중간전달체를 거쳐 페레독신까지 운반되는 것을 비순환적 전자전달이라고 하며, 이와 달리 전자가 명반응 Ⅰ안에서만 운반되는 것을 순환적 전자전달이라고 한다.
ATP의 합성
ATP는 아데노신이인산(ADP)에 인산기가 하나 더 붙는 인산화반응을 통해 만들어지는데 빛이 이 반응에 대한 에너지를 제공하기 때문에 이 반응을 광인산화(光燐酸化)라고 한다.
엽록체와 미토콘드리아에서의 ATP합성이 전자전달과 관련되어 있다는 이론을 처음으로 제시한 사람은 영국의 생화학자 피터 미첼이다. 가장 널리 인정받고 있는 이 화학삼투이론(化學渗透理論)의 핵심은 온전한 라멜라를 가로지른 수소 이온(H+, 양성자)의 농도기울기와 전위차의 형성이다.
여기서 수소 이온의 농도기울기와 전위차에 의해 저장된 퍼텐셜 에너지는 ADP와 무기인산(Pi)이 결합하여 ATP와 물을 생성하는 반응에 이용된다. 수소 이온의 농도기울기는 몇 가지 경로로 이루어진다. 명반응 Ⅱ에서 물이 광분해되면서 플라스토퀴논은 틸라코이드 바깥쪽에서 수소 이온을 받아 환원된 다음 시토크롬-철-황복합체에 전자를 전달하면서 틸라코이드 안쪽에 수소이온이 쌓이게 되므로 라멜라를 가로질러 수소 이온의 농도기울기를 형성한다.
또한 명반응 동안 음 전하를 띤 입자들이 라멜라 바깥으로 이동하기 때문에 라멜라를 가로질러 전위차가 생긴다. ADP와 Pi로부터 ATP가 생성되는 반응은 틸라코이드막 안쪽에 쌓인 H+가 이 막에 있는 ATP 분해효소복합체(ATPase complex)를 통과하면서 이루어진다.
탄소고정과 환원반응
탄소 경로의 발견
탄소동화의 경로를 밝히는 데는 탄소(C)와 인(P)의 동위원소인 14C와 32P가 사용되었다(→ 카본-14, 인-32). 자연상태의 CO2 대신 14CO2를 식물에 공급해 주었을 때도 똑같이 광합성을 하는데 이때 동화된 산물은 방사성 동위원소인 14C를 달고 있기 때문에 추적할 수가 있다.
초기 연구에서, 빛을 차단한 어두운 상태에서도 14CO2를 동화한 방사성 산물을 발견할 수 있었기 때문에 탄소고정 반응을 '암반응'이라고 했던 것이다. 탄소고정과 환원경로는, 이 연구로 노벨상을 받은 미국의 생화학자 멜빈 캘빈이 밝혔다.
캘빈은 14CO2가 공급된 상태에서 광합성을 한 식물의 생성물을 분석하여 여기에 3탄소화합물인 3-포스포글리세르산(PGA)·인산화 당·아미노산·설탕·카르복시산 등이 포함되어 있음을 알아냈다.
그러나 2초 동안만 광합성을 하고 멈추었을 때는 방사성을 띤 생성물이 대부분 PGA라는 것을 발견하여 이것이 첫번째 탄소고정 산물이라는 것을 알아냈다.
또한 32P를 지닌 무기인산을 이용해 5탄당인산환원회로(reductive pentose phosphate cycle:RPP cycle)라고 하는 탄소고정과 환원경로를 밝혔다(→ 포스포글루콘산 경로). 이러한 경로의 각 단계는 실험실에서 분리한 효소들에 의해 암조건에서 일어날 수 있으며, 이중 어떤 단계에서는 명반응에서 생긴 NADPH와 ATP가 이용된다.
5탄당인산 환원회로
탄소가 고정·환원·사용되는 RPP 회로에서는 중간대사물인 인산화당이 생성된다(그림 3). RPP회로가 완전히 한 번 도는 동안 3분자의 이산화탄소가 사용되어 3탄소화합물인 글리세르알데히드-3-인산(Gal 3P) 1분자가 생성된다.
이 3탄당 인산은 엽록체 밖으로 배출되거나 안에서 녹말로 바뀐다(점선). RPP 회로가 밤에도 작동한다면 쓸데없이 ATP를 소모하는 것이므로 밤에는 회로의 효소들이 작용하지 않는 것이 식물에 유리하다. 환원된 페레독신, 여러 가지 산, Pi, 마그네슘 이온 등의 수용성 요소들이 RPP 회로의 효소활성을 조절한다.
탄소환원의 산물
녹색식물에서 Gal 3P는 엽록체에서 세포질로 나와 설탕을 형성한다.
그밖의 주요산물로는 알라닌·글루탐산·아스파르트산과 같은 1차아미노산을 들 수 있다. 또한 지질, 색소의 합성에도 Gal 3P가 이용된다. 엽록체 안에서 녹말의 합성과 축적은 탄소고정이 필요이상으로 진행될 때 일어난다.
광호흡
덥거나 빛이 강하고 물이 부족할 때는 광호흡이 일어나기 때문에 RPP의 생산성이 떨어진다.
이 과정은 광합성 동안에 리불로오스-1,5-이인산(Ribulose 1,5-biphosphate:RuBP) 이 산소분자(O2)에 의해 산화되는 것을 시작으로 당이 이산화탄소로 바뀌기 때문에 광호흡이라고 한다. RuBP의 산화를 통해 1분자의 PGA와 2탄소인 포스포글리콜산 1분자가 생성되는데 이 산은 부분적으로 이산화탄소로 바뀐다.
이 과정은 식물세포 안의 엽록체·퍼옥시솜·미토콘드리아와 같은 세포 소기관 사이에서 일어나는 서로 연결된 여러 반응으로 이루어져 있다.
광호흡이 일어나는 이유는 산소분자에 의한 RuBP의 산화와 CO2에 의한 카르복시화 과정을 같은 효소가 촉매함으로써 이 두 과정이 서로 경쟁하기 때문이다.
4탄소(C4)산을 통한 탄소고정
열대에서 진화해왔을 것으로 생각되는 사탕무·옥수수 같은 식물들은 광호흡을 막을 수 있다.
탄소고정은 엽육세포에서 시작하지만 RPP회로에 의한 탄소환원은 실제로 관다발 근처에 있는 유관속초(有管束鞘) 세포에서 일어난다. 이런 식물에서는 이산화탄소가 4탄소산인 말산(malate)에 고정되어 유관속초 세포로 이동하여 세포 안의 이산화탄소 농도를 높여주기 때문에 광호흡이 줄어들어 에너지 손실이 적고 생산량이 높다.
광합성에 영향을 주는 요인
개요
육상식물의 광합성에 가장 직접적으로 영향을 미치는 환경요인으로는 빛의 양, 온도, 이산화탄소, 물의 공급, 무기물의 이용 가능성 등을 들 수 있는데 이러한 요인들은 건강상태·성숙도·생식시기 여부와 같은 생리적 조건이나 식물의 종(種)에 따라서도 달라진다.
빛의 세기와 온도
온도는 적절하지만 빛의 세기가 낮은 조건에서 광합성이 일어날 경우 광합성은 빛의 세기가 증가함에 따라 증가하다가 어느 정도 이상의 강한 빛이 되면 빛의 포화[光飽和]가 일어나 더 증가하지 않는다.
따라서 광합성은 광포화가 일어나기 전까지는 광화학 반응의 속도에 의존하지만 포화가 일어난 뒤부터는 빛과 관계없이 온도에 의존하게 되고 빛과 상관없는 암반응의 화학반응이 광합성률의 제한요소가 된다. 또한 대부분의 육상식물에서는 빛의 세기와 온도가 모두 높을 때는 광호흡 현상이 일어나 광합성의 증가를 제한한다.
이산화탄소
광합성의 암반응은 이산화탄소가 유기물로 바뀌는 화학반응이기 때문에 이 반응의 속도는 공기 중의 이산화탄소 농도에 따라 달라진다.
물
식물체 안의 물은 증산작용을 통해 잎의 기공에서 대기로 증발한다.
기공은 잎의 표피에 있는 작은 구멍인데 이것을 통해 식물체 안으로 이산화탄소가 들어오고 산소와 물이 대기로 나간다. 덥고 건조한 기후조건에서 식물은 물을 보존하기 위해 기공을 닫지만 이렇게 될 경우 이산화탄소가 들어오지 못해 광합성률이 낮아지며 불필요한 광호흡만 증가한다.
무기물
식물이 잘 자라고 광합성을 최대로 유지하기 위해서는 몇 가지 무기물이 반드시 필요하다.
질산·암모니아·황산염·인산·철·마그네슘·칼륨은 많은 양이 필요하며, 그밖에 아주 적은 양의 망간·구리·염소 등이 필요하다.
내부 요인
각 식물종들은 효소 활성을 조절할 수 있는 복잡한 조절기구를 통해서 여러 환경요인들에 적응을 하는데, 광합성 역시 이러한 기구를 통해 식물체 전체의 필요에 따라 조절된다.
중요성과 연구전망
전세계적으로 인구와 인간활동의 폭발적인 증가에 따라 경작지와 삼림이 빠른 속도로 줄어들고 있다. 따라서 제한된 땅·물·시간에서 더 많은 생산을 해야 하는 상황을 맞게 되었다. 과거에는 이러한 문제를 주로 비료의 개선과 식물교배를 통해 해결해왔으나, 1970년대 이후부터는 유전공학기술을 사용하여 기본적인 농업생산성 증대를 꾀하고 있다.
식물의 생산성은 태양에너지가 광합성과정을 통해 얼마나 효율적으로 화학에너지로 저장되느냐에 달려 있는데, 최근에는 분자생물학적 방법을 이용하여 빛에너지와 전자전달에 관여하는 단백질들을 변형시킴으로써 이 과정의 자세한 단계들이 밝혀졌다. 이로써 식물의 생산성을 증대시킴은 물론 물이 부족하거나 염분이 많은 곳에서도 경작을 할 수 있게 되었고, 제초제에 대한 저항성 식물도 개발하여 선택적인 제초효과도 얻을 수 있게 되었다.
| |||||||||||
| |||||||||||



물은 생명이 존재할 가능성을 나타내지만, 산소는 그 가능성이 실현되었음을 뜻한다. 오직 생물만이 독립적으로 존재하는 산소를 많든 적든 공기 중에 내놓을 수 있기 때문이다.
- 닉 레인, ‘산소’에서
게놈 데이터가 만능은 아니지만 많은 경우 결정적인 역할을 한다. 해묵은 논쟁을 끝내기도 하고 이전엔 생각해보지도 못했던 사실을 드러내 새로운 시각을 갖게 하기도 한다.
예를 들어 네안데르탈인과 현생인류의 혼혈 여부는 수십 년 동안 피가 섞이지 않았을 것이라는 진영이 우세한 채 논란이 됐지만 2010년 네안데르탈인의 핵 게놈이 해독되면서 단번에 해결됐다. 즉 아프리카를 제외한 지역에 사는 사람들은 네안데르탈인의 피가 섞인 것으로 나오면서 현생인류의 이동 시나리오와도 맞아떨어지자 반대 진영에서 더 이상 할 말이 없어진 것이다. 새끼손가락 뼈 한 마디에서 추출한 DNA에서 고품질의 게놈이 해독돼 수만 년 전 아시아에 데니소바인이라는 미지의 인류가 살았다는 사실이 드러나기도 했다.
미생물 분야는 게놈 데이터로 연구 패러다임이 바뀐 지 꽤 됐다. 예전에는 일단 배양이 돼야 미생물을 연구할 수 있었지만 세포 하나만 있어도 게놈을 분석할 수 있는 지금은 메타게놈, 즉 시료에 있는 모든 생명체의 게놈을 통째로 분석하는 게 일상이다. 그 결과 장내미생물 연구를 비롯해 많은 분야에서 놀라운 사실들이 많이 밝혀졌다.

● 23억 년 전 산소 존재감 드러내
학술지 ‘사이언스’ 3월 31일자에는 ‘시아노박테리아(남세균)에서 산소발생 광합성과 산소호흡의 기원에 대하여’라는 다소 거창한 제목의 논문이 실렸다. 배양이 안 되는 시아노박테리아 41종의 게놈을 분석한 결과 광합성과 호흡의 기원까지 추측할 수 있게 됐다는 내용이다. 특히 산소발생 광합성은 시아노박테리아가 가장 먼저 시작했으므로 ‘지구에서 산소발생 광합성의 기원에 대하여’인 셈이다.
이 시점에서 몇몇 독자들은 광합성 앞에 굳이 ‘산소발생’이란 수식어를 쓸 필요가 있는지 의아할 것이다. 광합성의 정의를 ‘빛에너지로 물분자에서 전자를 뽑아 이산화탄소를 당(유기물)으로 환원시키고 노폐물로 산소를 내보내는 과정’으로 알고 있는 경우인데 사실 이게 산소발생 광합성(oxygenic photosynthesis)의 정의다. 그러나 광합성에 필요한 전자를 꼭 물분자에서 얻을 필요는 없고 실제 제1철이온(Fe2+)이나 황화수소(H2S) 같은 다른 물질에서 얻는 광합성 과정이 널리 쓰이고 있다. 이 경우 물론 폐기물로 산소가 나오지 않는다. 이를 산소비발생 광합성이라고 부른다.
영국의 생화학자이자 저술가인 닉 레인은 2002년 ‘산소’라는 대단한 책을 펴냈는데 2004년 번역서가 나왔지만 출판사가 문을 닫으며 절판됐다가 얼마 전 재출간됐다. 이 책을 보면 오늘날 지구가 이처럼 아름다운 행성이 된 건 시아노박테리아 덕분이라는 생각이 절로 든다. 시아노박테리아 등장 전까지 지구에 바다는 있었지만 육지는 황무지였고 대기에는 산소가 거의 없었다. 그 결과 산소를 필요로 하지 않는 미생물들만이 번성하고 있었다.
그런데 여러 지질학 증거에 따르면 23억 년 전을 전후해 대기 중 산소의 농도가 눈에 띠게 늘어나 현재의 5~18% 수준이 됐고 이렇게 10억 년 이상을 보내다가 약 8억 년 전부터 산소 농도가 높아지기 시작해 대략 6억 년 전 현재 수준에 이른 것으로 보인다. 따라서 시아노박테리아는 늦어도 23억 년 전에는 지구에 모습을 드러냈다는 말이다.
2014년 학술지 ‘네이처’에 실린 한 리뷰논문을 보면 산소발생 광합성이 시작된 시점이 여전히 불확실해 학자에 따라 멀게는 38억 년 전에서 가깝게는 23억5000만 년 전까지로 무려 15억 년 가까이 차이가 난다는 구절이 나온다. 지구의 나이가 45억 년이므로 거의 3분의 1만큼이 오차인 셈이다. 닉 레인의 책도 그렇고 필자도 38억 년 쪽(또는 37억 년, 35억 년)으로 알고 있었는데 어느새 훨씬 짧게 추정하는 가설이 꽤 입지를 다졌나보다.
산소발생 광합성이 35억~38억 년 전 시작됐다고 주장하는 입장은 지질 데이터나 화석을 근거로 든다. 즉 동위원소비나 특정 화합물의 존재, 스트로마톨라이트 같은 화석이 증거다. 예를 들어 호주 서부의 35억 년 전 지층에서 발견된 스트로마톨라이트를 시아노박테리아의 화석으로 보고 이때 이미 산소발생 광합성이 일어났다는 설명은 오늘날 정설로 받아들여지고 있다. 산소발생 광합성 시작과 대기 중 산소 농도 증가 시작 시점이 10억 년 이상 차이가 나는 건 그사이 발생한 산소가 호흡으로 재순환되거나 바닷물에 녹고 암석을 산화시키는데 소모됐기 때문이라고 설명한다.
그런데 21세기 들어 이런 해석에 회의적인 입장이 늘고 있다. 지질학적 증거도 얼마든지 다른 식으로 해석할 수 있고 어떤 경우는 시료가 오염된 것으로 보이며(후대 지층과 섞임) 그 옛날 스트로마톨라이트를 오늘날 스트로마톨라이트와 동일시해 시아노박테리아 덩어리라고 보는 것도 무리다. 즉 시아노박테리아는 고사하고 미생물인지 아닌지도 불확실하다는 것이다. 시아노박테리아가 분명한 화석 가운데 가장 오래된 건 ‘불과’ 19억 년 됐다.
이런 와중에 시아노박테리아에 대한 전혀 생각지도 못한 게놈 데이터가 나오면서 회의적인 입장이 힘을 얻고 있다. 이번에 ‘사이언스’에 실린 논문이 결정판으로 이에 따르면 빨라야 25년 전이나 26억 년 전 산소발생 광합성이 시작한 것으로 나온다.
● 깜깜한 장 속에 시아노박테리아 산다고?
일은 엉뚱한 데서 시작됐다. 2005년 동물의 장내미생물에 대한 메타게놈 연구에서 시아노박테리아에 속하는 것으로 보이는 유전자의 데이터가 좀 나왔다. 빛이 전혀 들어가지 않는 장 속에 광합성을 하는 박테리아의 유전자가 있다는 뜻밖의 결과였지만 박테리아 사이에는 유전자수평이동도 있으므로 아주 불가능한 일은 아니다.
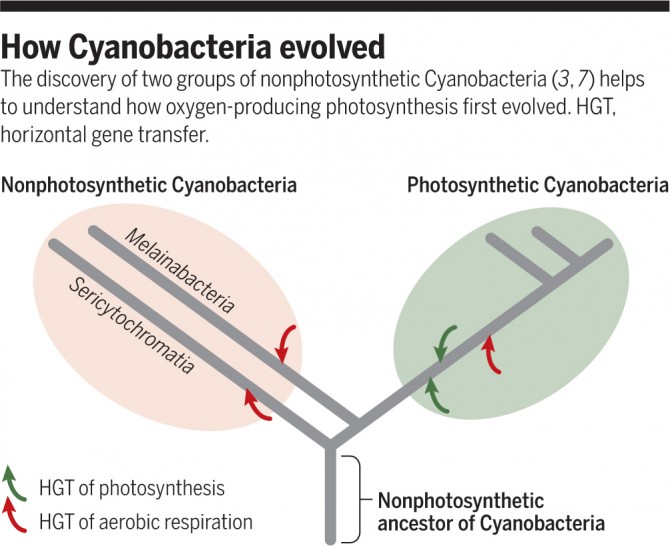
그러나 2013년 사람의 장과 지하수 같은 깜깜한 곳에서 얻은 시료에서 시아노박테리아의 게놈이 해독되면서 이야기는 새로운 국면으로 접어들었다. 즉 우리가 지금까지 알고 있었던 시아노박테리아는 실제 그 일부였던 셈이다. 게놈분석 결과 깜깜한 곳에서 살고 있는 시아노박테리아에는 예상대로 광합성을 하는 유전자 자체가 없었다. 연구자들은 이 그룹을 멜라이나박테리아(Melainabacteria)라고 부르며 기존 산소발생 광합성을 하는 시아노박테리아에도 옥시포토박테리아(Oxyphotobacteria)라는 이름을 붙였다. 그러나 멜라이나박테리아가 원래는 광합성을 할 수 있었지만 어쩌다 빛이 들어오지 않는 환경에 놓여 필요 없어진 유전자들을 잃어버렸을 가능성도 있었다.
그런데 이번에 41종의 게놈을 분석한 결과 시아노박테리아 진화의 진면목이 밝혀진 것이다. 먼저 앞의 두 그룹에 속하지 않는 종들이 여럿 확인되면서 세리사이토크로마티아(sericytochromatia)라는 새로운 그룹이 더해졌다. 한편 세리사이토크로마티아에도 광합성관련 유전자가 없다. 그리고 세 그룹의 계통을 비교한 결과 공통조상에서 세리사이토크로마티아가 먼저 떨어져 나가고 그 뒤 옥시포토박테리아와 멜라이나박테리아가 갈라졌다. 연구자들은 분자시계, 즉 DNA 염기의 변이 정도를 비교한 결과 이들이 갈라진 시점이 대략 25년 전이나 26억 년 전으로 추정했다.
즉 오늘날 시아노박테리아의 공통조상은 광합성을 하지 않았고 세 그룹이 갈라진 뒤 옥시포토박테리아의 조상이 어느 시점에서 광합성 능력을 얻었다는 것이다. 연구자들은 박테리아 사이의 유전자수평이동을 통해 옥시포토박테리아가 산소비발생 광합성 유전자를 얻었고 그 뒤 물을 분해하는 산소발생복합체를 진화시키면서 산소발생 광합성을 하는 유일한 박테리아가 됐다고 추정했다. 그 뒤 이 박테리아의 한 종이 진핵생물에 잡아먹혀 소화되는 대신 세포 내에 자리를 잡으며 엽록체가 됐고 그 결과 조류(algae)와 식물이 나왔다.
한편 이들 세 그룹의 산소호흡, 즉 당을 산소로 산화해 에너지를 얻는 세포호흡에 관여하는 유전자를 분석하자 각자 계열이 다른 것으로 드러났다. 즉 산소호흡은 세 그룹이 나뉜 뒤 독자적으로 진화했다는 말이다(역시 유전자수평이동을 통해 관련 유전자를 획득했을 것이다). 산소호흡은 외부의 산소농도가 어느 정도 높는 환경에서 쓸모가 있으므로 시기적으로도 말이 된다.
이번 연구결과가 산소발생 광합성 시기 논쟁에 어느 정도 영향을 미칠지 아직 예단하기는 이르지만 기존 주류 입장(38억~35억 년 전)에는 꽤 큰 타격을 줄 전망이다. 35억 년 전 스트로마톨라이트가 설사 시아노박테리아로 입증된다고 할지라도 그게 산소발생 광합성의 증거는 될 수 없기 때문이다.
'숲과의 對話 > 숲 일반' 카테고리의 다른 글
| MBTI 16가지 성격유형과 검사 (0) | 2017.04.12 |
|---|---|
| 산림해충 (0) | 2017.04.09 |
| 산림토양 (0) | 2017.04.09 |
| 줄기의 구조와 형태 (0) | 2017.04.05 |
| 잎의 구조와 형태 (0) | 2017.04.05 |